单细胞文章解析(2):拟南芥茎尖单细胞分析
A single-cell analysis of the Arabidopsis vegetative shoot apex
一、文章概述
在植物的整个生命周期中,茎顶端分生组织允许新的气生结构的反复形成。我们使用单细胞RNA测序在转录组水平上定义拟南芥茎尖的细胞分类。我们发现拟南芥茎尖由高度异质性的细胞组成,它们可以被划分为7个主要的群体,其中包含23个转录组水平上较为差异的细胞类群。作者描述了表皮细胞、维管组织和叶肉细胞的细胞周期连续性和发育轨迹,并推断出决定细胞命运的转录因子和基因的表达特征。最后,作者对茎尖和根尖细胞群体的综合分析进一步揭示了表皮和维管组织的相似性与差异性。该篇文章的研究结果为探究植物细胞分裂和分化的基本原理提供了一个有价值的资源。
二、研究背景
除下胚轴和子叶外,植株地上部分均由茎尖分生组织(SAM)产生。被子植物的SAM由具有特定结构特征的小圆顶细胞组成。具研究SAM的不同层或区域调控着干细胞稳态和叶原基初始化。在克隆研究的基础上,拟南芥SAM可分为三个层。L1层的细胞可分化形成芽、叶和花的表皮,而L2层形成中胚层组织和生殖细胞,L3层形成维管组织和髓。根据植物形态学和组织学研究,拟南芥的SAM也可以分为不同的区域。中央区域(CZ)是干细胞聚集的地方。外周区(PZ),这里的细胞分裂比CZ更频繁,负责器官的初始化,以及肋骨分生组织(RM),它产生茎轴的中心组织。
植物干细胞的生态位是在胚胎发育过程中确定的。在动物中,干细胞聚集在干细胞生态位(SCN)中,SCN产生信号,调节干细胞的分裂和分化。在茎尖组织中,维持干细胞的数量需要在组织中心(OC)区域表达同源结构域蛋白WUSCHEL (WUS)以及在中央区域(CZ)表达CLV3。茎尖干细胞经过几轮分裂,最终分化并产生不同类型的器官,如叶和花。
尽管前人已经对经尖分生组织进行了大量的研究,但是对茎尖上每个细胞的特性仍缺乏全面的了解,因此,这限制了我们探究分生组织功能和描述分生组织细胞分化轨迹。此前的分析研究使用报告基因纯化细胞群,这是一种低估细胞复杂性的方法。与bulk RNA测序相比,单细胞RNA测序(scRNA-seq)实现了更高的分辨率,能够表征植物中不同细胞群的亚群的许多特性。例如,几个scRNA-seq研究表明,拟南芥根尖细胞在转录组中是高度异质性的。不同发育阶段个体细胞的空间分布和时间顺序揭示了根系发育的连续分化轨迹。
三、结果解读
3.1 拟南芥茎尖细胞图谱的绘制
3.1.1 材料获取与聚类分群
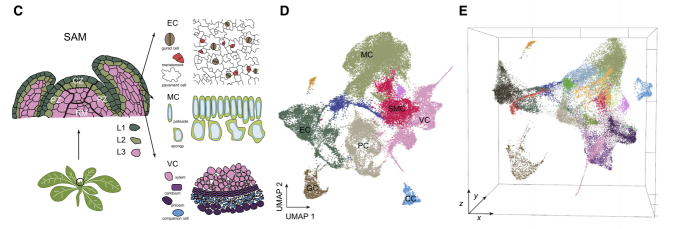
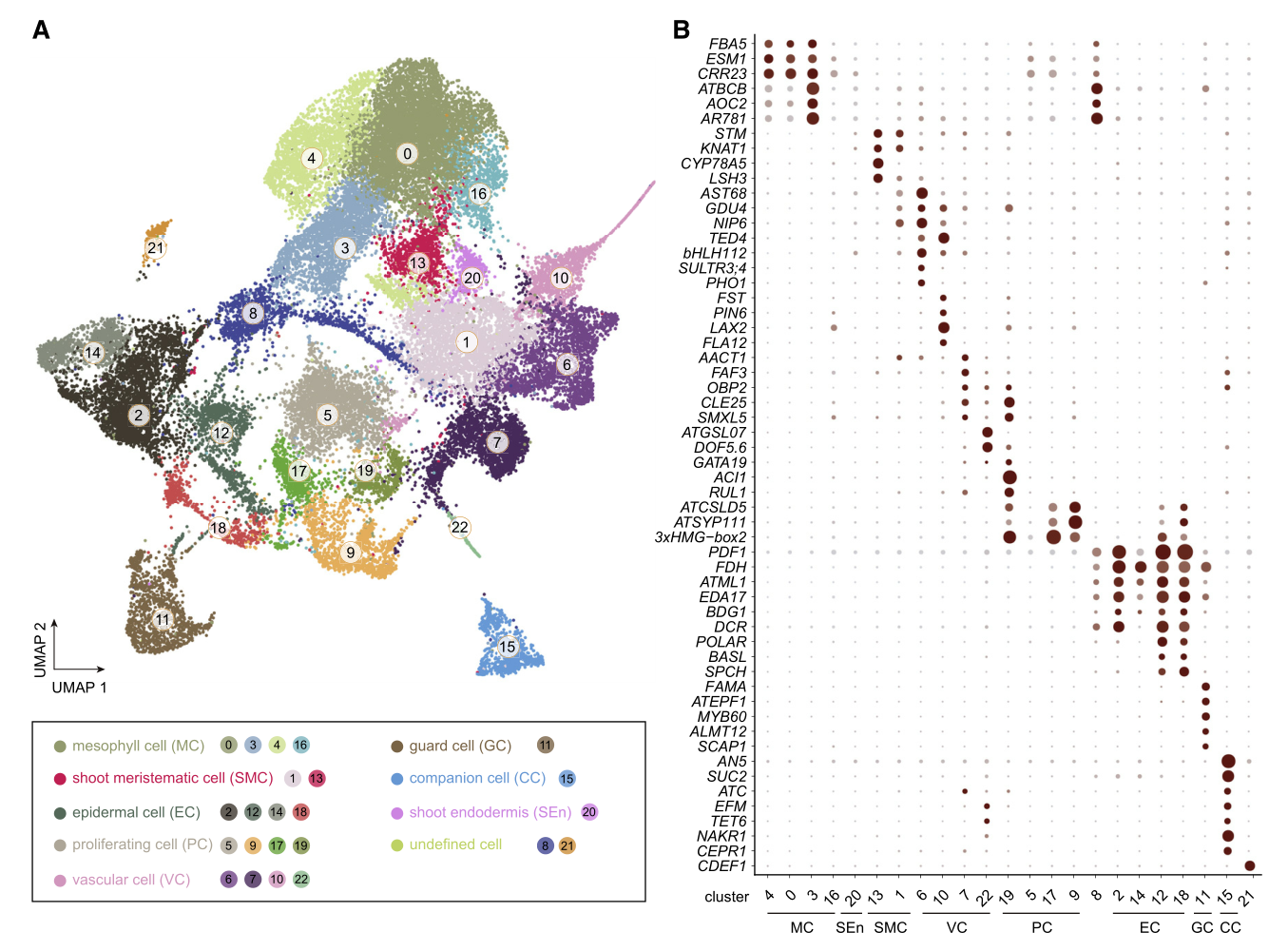
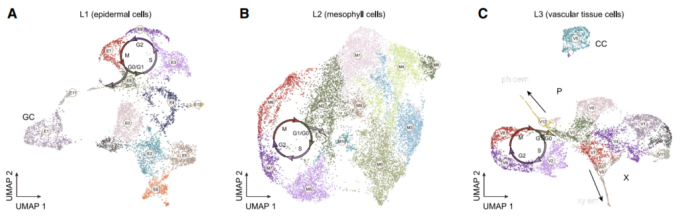
由于拟南芥茎尖非常的微小由SAM和叶原基组成,从茎尖分离出分生组织也就十分的困难。作者获取了500个茎尖,从而增加了样品中茎尖分生组织细胞(SAM)的数量。为了区分SAM与叶细胞类群,作者同样采摘了18天的幼叶。茎尖和叶片材料分别进行消化获得原生质体,然后通过10X Genomics平台进行单细胞测序。茎尖样品做了两次生物学重复。 在后续的分群聚类过程中,作者还消除了原生质体制备过程和细胞周期因素对分群的影响。最终,作者从茎尖和叶片的36643个细胞中鉴定出23个细胞类群,这些细胞类群最后被归为7个较为大的类群,分别是:叶肉细胞(mesophyll cell,MC)、茎尖分生组织细胞(shoot meristematic cell,SMC)、维管组织细胞(vascular cell,VC)、表皮细胞(epidermal cell,EC)、增殖细胞(proliferating cell,PC)、保卫细胞(guard cell,GC)和伴随细胞(companion cell,CC)。
作者研究的是SAM为什么还要加上叶片细胞?
3.1.2 各类群的鉴定

叶肉细胞类群(Clusters 0, 3, 4, and 16)
光合作用基因CHLORORESPIRATORY REDUCTION23 (CRR23), *RUBISCO SMALL SUBUNIT 2B (RBCS2B)和PHOTOSYNTHETIC NDH SUBCOMPLEX L1 (PNSL1)*这三个基因主要在这几个类群中表达。通过检测富集到的marker基因AT1G13650的启动子活性,该类群的推断得到了进一步证实。
表皮细胞类群(Clusters 2, 12, 14, and 18)
在EC细胞类群中检测到表皮组织特异性表达的基因MERISTEM LAYER1 (ATML1), PROTODERMAL FACTOR1 (PDF1)和FIDDLEHEAD (FDH)。
茎尖分生组织细胞(Clusters 1, 13)
建立和维持拟南芥SAM所需的基因SHOOT MERISTEMLESS (STM)以及同源结构域基因KNAT1, KNAT2 , KNAT6在这两个类群中富集表达。
增殖细胞类群(Clusters 5, 9, 17, and 19)
与细胞周期相关的三个基因如*HISTONE H4 (HIS4)、CYCLIN-DEPENDENT KINASE B2;1 (CDKB2;1)和CY-CLIN A1;1 (CYCA1;1)*在这四个类群中富集表达。
维管组织细胞类群(Clusters 6, 7, 10, and 22)
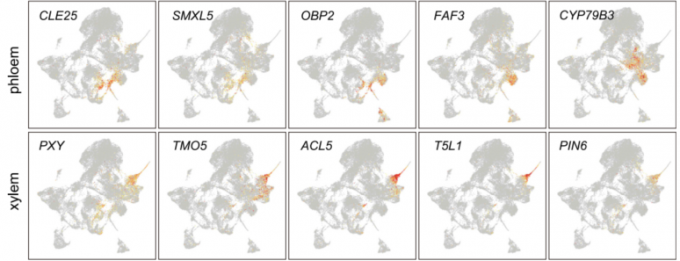
VC群体由4个细胞类群(6、7、10、22)组成,其中木质部和韧皮部分化相关基因富集表达。例如,韧皮部基因*SMAX1-LIKE5 (SMXL5)和木质部基因PHLOEM INTERCALATED WITH XYLEM (PXY)*的转录本分别在第7和第10细胞类群中显著富集。
保卫细胞类群(Clusters 11)
*FAMA (FMA)、EPF1 (EPIDERMAL PATTERNING FACTOR1)*等GC的marker基因在Cluster 11细胞类群中特异性富集。
伴随细胞类群(Cluster 15)
*PHLOEM PROTEIN2-A1 (PP2-A1) and SUCROSE-PROTON SYMPORTER2 (SUC2)*等CC的marker基因在Cluster 15细胞类群中特异性富集。
内皮层细胞类群(Cluster 20)
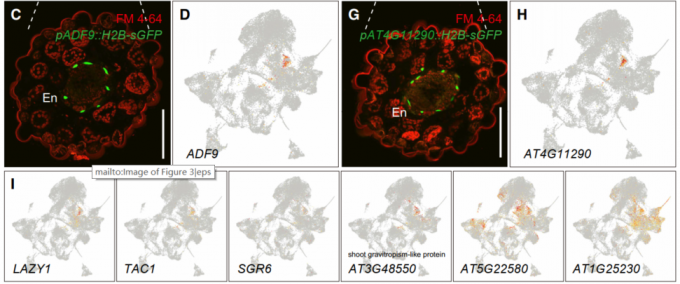
Cluster 20富集了参与芽向地性反应的基因,表明存在一个独特的细胞群体,负责在芽中感知重力。对该类群中的两个marker基因(ACTIN DEPOLYMERIZING FACTOR9和AT4G11290)的启动子报告分析表明,Cluster 20细胞类群属于芽内皮层细胞类群。
未知的细胞类群(Cluster 8, 21)
Cluster 8, 21由于缺乏已知的标记基因而无法得到注释。此外,RNA荧光原位杂交检测特异性基因同样没有产生可靠的信号。GO富集分析显示,该类群被标注为生物过程类别“应激反应”的基因占主导地位。
3.2 控制芽枝角度相关基因的鉴定
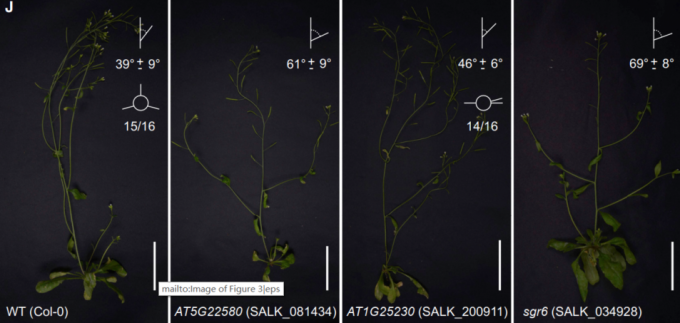
在细胞类群的分群聚类注释的过程中,文章鉴定出每个类群中特异表达或者富集表达的基因,那么这些基因对植物茎的生理发育又有着哪些影响,作者希望能通过反向遗传学来试着挖掘出一些有价值的信息。本文作者重点观察了了Cluster 20这个群体,之前对该群体的marker gene进行功能富集分析发现,该类群中的基因主要富集于茎对重力响应的功能。之前研究发现,LAZY1, TILLER ANGLE CONTROL1 (TAC1), and SHOOT GRAVITROPISM6 (SGR6)这些基因都与茎的重力响应相关。在该类群中作者选择了48个marker基因,从种子库中心获得相应的纯合子的T-DNA插入突变体。最终,作者发现AT5G22580和AT1G25230这两个基因的突变体的表型与sgr6植株的表型一致都显示出大分枝的表型。
3.3 表皮细胞、叶肉细胞与维管细胞中的细胞周期特性
植物发育的关键因素之一是在茎尖通过细胞分裂和分化形成器官。Transit-amplifying细胞是一群未分化的细胞,被认为是干细胞与已分化细胞的中间状态。为了了解表皮、叶肉和维管组织的发育轨迹,需要识别和表征相应的早期分裂细胞,分化中的细胞和已经分化的细胞。
3.3.1 表皮细胞类群
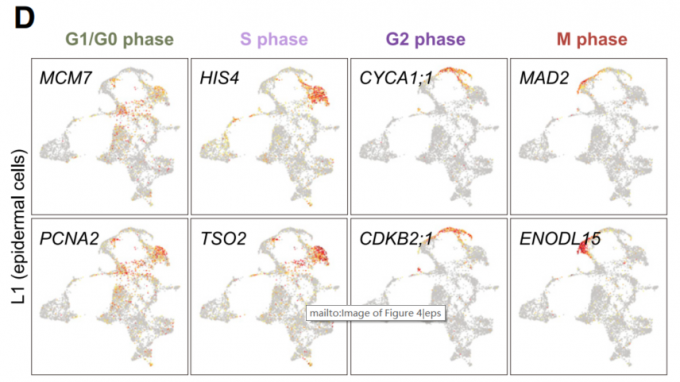
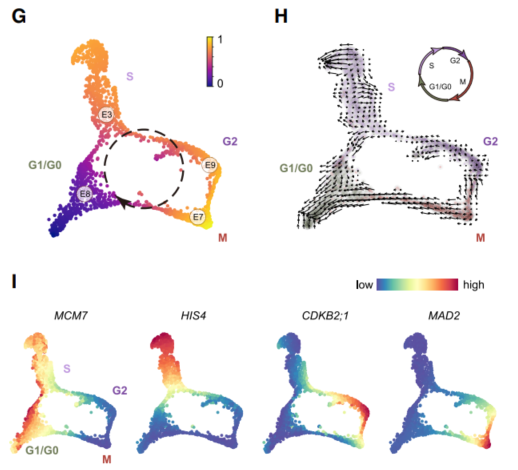
首先,对所有的表皮细胞重新进行聚类分群,最终得到13个亚群(E0~E13),E3,E7,E8,E9四个亚群形成一个闭环结构。
其次,(1)由于DNA合成基因HIS4 和 TSO2在E3中富集表达,因此E3被识别为S期细胞类群;(2)CDKB2;1 and CYCA1;1基因在E9中富集表达,因此E9被识别为G2期细胞类群;(3)M期细胞类群由E7组成,该亚群中富集表达*EARLY NODULIN-LIKE PROTEIN15 (ENOD15)和MITOTIC ARREST-DEFICIENT2 (MAD2)这两个M期标志性基因。(4)最后,由于MINICHROMSOME MAINTENANCE7 (MCM7)、PROLIFERATING CELL NUCLEAR ANTIGEN2 (PCNA2)、CYCLIN D3;2 (CYCD3;2)*和核糖体蛋白编码基因如核糖体蛋白L24e (RPL24)和核糖体蛋白S6 (RPS6)的高表达,将E8亚细胞群划分为G0或G1期。
3.3.2 叶肉细胞类群与维管细胞类群
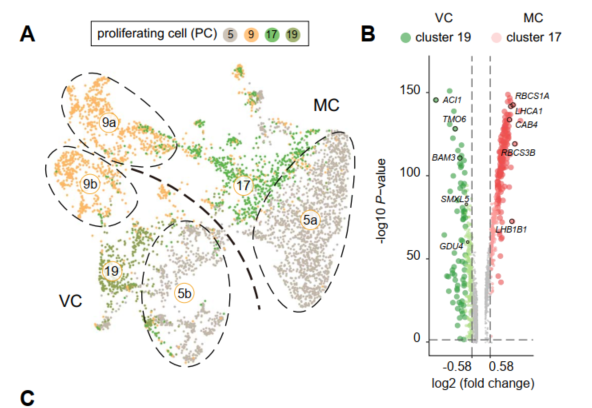
首先,依照前面的分析思路作者解析叶肉细胞与维管细胞群体中的细胞分裂活动。但是,在MC和VC群体中并没有观察到与EC群体相似的细胞周期分布。作者推断这两个群体中与细胞周期相关的群体可能分布在增殖细胞群体中。通过对PC细胞类群进行重新群类分群,Cluster5和Cluster9被分成了两个亚群(Clusters 5a and 5b; Clusters 9a and 9b)。
由于与光合作用相关的基因RBCS2B和PNSL1在Clusters 17, 5a, and 9a中富集分布,而维管组织的marker基因SMXL5和*GLUTAMINE DUMPER4 (GDU4)*主要在clusters 19, 5b, and 9b中富集分布。这说明PC群体主要由两个分裂细胞群体组成,从而导致了叶肉细胞和维管组织的分化。通过对这两个群体重新分群聚类,最终作者挖掘出与表皮细胞类群中相似的细胞周期现象。
最后,基于EC、reMC和reVC群体中在相同细胞周期阶段表达的基因,创建了一个拟南芥基因核心目录,并记录其转录水平在细胞周期中周期性变化。
3.4 表皮细胞向气孔分化的研究
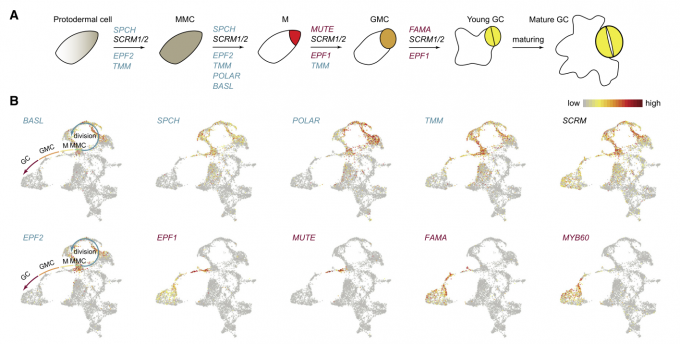
高度分化的表皮细胞是在叶片发育过程中由表皮原始细胞分化而来的。气孔是由分生组织母细胞通过细胞分裂和一系列分化而产生的。为了探究气孔细胞的生成过程,作者将表皮细胞类群中的E3, E7, E8, and E9归为细胞分裂亚群,将E1, E8, and E11归为分化亚群。
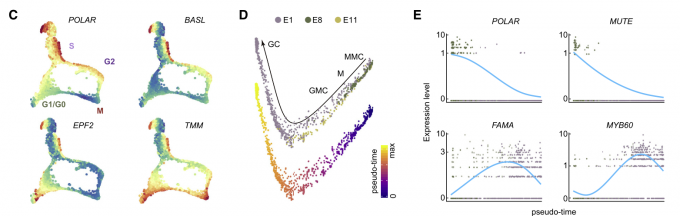
- 一些与气孔发育相关的早期基因如POLAR, SPCH, BASL, TMM,在分裂亚群中富集表达,特别是参与分生体母细胞不对称分裂的基因如POLAR和BASL在S期高表达。而抑制气孔分化的基因TMM和EPF2的表达量主要在M期或G0/G1期达到峰值。因此,这些基因的时间表达模式可能将其产物的积累与细胞的不对称分裂或保卫细胞分化所需的特定时间窗口联系起来。
- 通过拟时间分析,对EC群体E1、E8和E11聚类细胞进行排序,揭示了气孔的后期分化轨迹。与之前的报道一致,调控这些细胞命运转变的关键转录因子,包括MUTE、FMA和MYB60,沿着细胞分化轨迹依次表达。
3.5 维管组织的分化轨迹
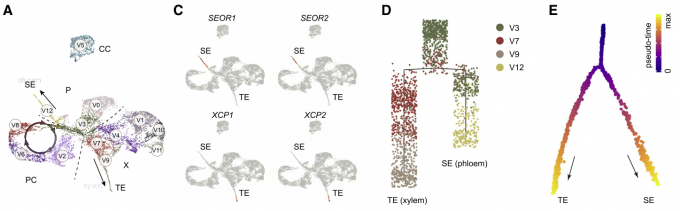
维管分生组织中的干细胞,也称为原形成层或形成层,通过不对称的周壁细胞分裂,连续产生两个传导组织,木质部和韧皮部。同样,作者把4.3中的重新聚类的维管组织细胞(reVC)分为分裂亚群(V2, V3, V6, and V8)和分化亚群(V3, V7, V9, and V12)。通过RNA原位杂交实验的验证,韧皮部由V0, V5, and V12细胞类群组成,V1, V4, V7, and V9-11组成了木质部。V12被鉴定为韧皮部的筛管细胞类群,V9鉴定为木质部的导管细胞类群,因为SEOR1,SEOR2,XCP1,XCP2基因在这些类群中富集分布。
在拟时序分析中,作者同样鉴定出了TEs (xylem)和SEs (phloem)两条分化轨迹。与植物激素生长素促进木质部分化的作用相一致,在木质部分化轨迹上与生长素通路相关的基因,如MONOPTEROS (MP), IAA12, LIKE AUXIN RESISTANT2 (LAX2), and PIN-FORMED6 (PIN6)都富集表达。GO富集分析进一步表明,该分支高度富集了生长素反应相关的生物学过程。同样,SMXL5, ALTERED PHLOEM DEVELOPMENT (APL), HIGH CAMBIAL ACTIVITY2 (HAC2/DOF5.6), and SEOR1等基因在韧皮部的拟时序轨迹上富集表达。
综上所述,这些数据表明,scRNA-seq使我们能够揭示维管组织中新的细胞类型,并揭示木质部和韧皮部分化过程中隐藏的过渡细胞状态。
3.6 茎分生组织(SMC)特性分析
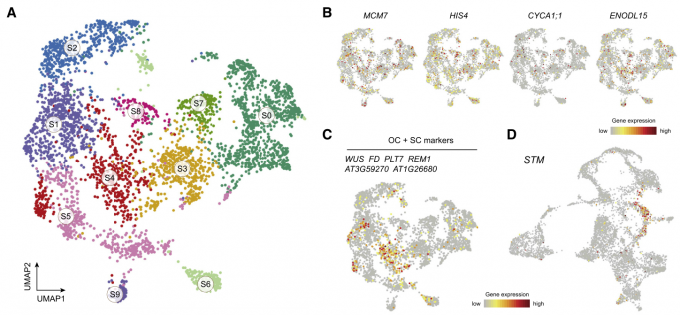
通过对茎分生组织细胞类群的重新聚类与分群,最终所有细胞被分为了10个亚群。细胞周期基因分析显示SMC转录组异质性不受细胞周期状态的影响。由于表达WUS和CLV3的细胞数量较少且表达水平较低,我们结合已知OC和CZ标记基因,包括PLETHORA7 (PLT7)、REPRODUCTIVE MERISTEM1 (REM1)、AT3G59270和AT1G26680,将簇S4命名为OC和CZ区。
众所周知,STM基因在SAM中高度表达,在叶原基中受到抑制。由于STM基因不仅在SMC类群中表达而且在EC类群中也表达,作者根据这一现象认为,分化中的SAM细胞可能分布在已经分化的细胞类群中,而不是SMC类群中。作者然后把所有表达STM的细胞进行新的聚类分群,最终得到了9个细胞亚群,W0到W8。依据marker基因对每个亚群进行了注释。
转录因子在决定细胞命运中起着关键作用。STM+细胞的基因共表达分析显示,STM与KNAT1高度共表达,但与正在分化的细胞类群中富集的基因共表达系数较低。
3.7 reMC(叶肉细胞类群)特性分析
除了与细胞周期相关的细胞亚群外,在reMC群体中还有7个不同的细胞亚群。遗憾的是,GO项分析和聚类特异性基因并没有识别出这些细胞亚群的任何功能。尽管人们普遍认为叶肉细胞由两种类型的细胞组成,即海绵细胞和栅栏细胞,但叶肉细胞的特性和功能比我们之前认为的要复杂得多。
3.8 单细胞分辨率下茎和根根尖组织的比较
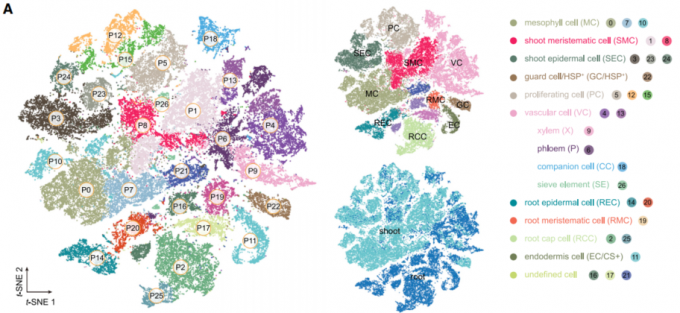
最后,作者尝试用单细胞分辨率分析茎尖和根尖之间的保守性和分化性。如图8A所示,结合根和茎尖的scRNA-seq数据集,包括60099个细胞,显示了27个细胞类群(P0到P26)。
- 大多数的细胞类群几乎都是由根或茎尖细胞组成。
- P22细胞群体主要由保卫细胞(GC)组成,27%的细胞来源于根。对P22类群中富集的基因的功能分析显示,*HEAT SHOCK PROTEIN (HSP)*基因的转录水平很高,这表明一些根细胞与GC具有转录组相似性,可能是因为它们对环境条件具有相同的高度敏感性质。因此,作者将P22标注为GC/HSP+。
- 茎尖和根尖组织共享6个维管组织细胞类群,这些共享的细胞被分为14个细胞亚群(l0到l12)。其中,l11和l12两个亚群分别由根尖细胞和茎尖细胞组成。
- 茎尖的表皮细胞类群(P3, P23, and P24)与根的表皮细胞类群(P14 and P20)并没有交集。但是作者发现,气生表皮的marker基因ATML1同样也在根尖的表皮细胞中表达。此外,一组参与表皮角层生物合成的基因,包括FIDDLEHEAD、FATTY ACID REDUCTASE1、GPAT4、GPAT8和3-KETOACYL-COA SYNTHASE20在茎和根的ATML1+细胞中共表达。这一结果与最近的发现一致,即在初生根尖和新生侧根中存在类角质层细胞壁结构,这也同样说明了ATML1在根表皮生物合成中发挥重要作用。综上所述,上述结果揭示了茎和根表皮之间以前未被认识到的相似性,并揭示了ATML1在根EC分化中的作用。
总结
- 茎尖细胞图谱的构建:茎尖和叶片的36643个细胞中鉴定出23个细胞类群,这些细胞类群最后被归为7个较为大的类群,分别是:叶肉细胞(mesophyll cell,MC)、茎尖分生组织细胞(shoot meristematic cell,SMC)、维管组织细胞(vascular cell,VC)、表皮细胞(epidermal cell,EC)、增殖细胞(proliferating cell,PC)、保卫细胞(guard cell,GC)和伴随细胞(companion cell,CC)。
- 细胞周期性研究:对表皮细胞类群,增殖细胞类群中的reMC和reVC两个类群的细胞周期性图谱进行探索与研究
- 细胞分化轨迹研究:(1)表皮细胞类群中气孔形成的分化轨迹;(2)维管组织细胞类群中,木质部与韧皮部的分化轨迹;(3)茎分生组织细胞向其他类型细胞分化过程中一些关键调控因子的鉴定。
- 单细胞分辨率下茎和根根尖组织的比较:(1)大部分细胞类群是相互独立的,两个组织中相似的细胞类群较少,主要集中在维管组织类群;(2)鉴定出ATML1基因在根和茎的表皮细胞中的功能。
单细胞文章解析(2):拟南芥茎尖单细胞分析

