单细胞文章解析(1):单细胞转录组图谱和染色质可及性揭示了水稻根的分化轨迹
文章:Single-cell transcriptome atlas and chromatin accessibility landscape reveal differentiation trajectories in the rice root
一、文章概述
根的发育依赖于分生组织的建立,分生组织产生不同的细胞类型,这些细胞类型在确定的时间和空间梯度上分化。对双子叶和单子叶根尖分生组织的发育轨迹和转录网络进行剖析,有助于理解双子叶和单子叶根尖分生组织的功能。在这里,我们提出了水稻胚根的单细胞RNA (scRNA)测序和染色质可及性调查。通过单个根尖细胞的时间分布,我们重建了表皮细胞和基底组织的连续发育轨迹,并阐明了这些细胞谱系中决定细胞命运的调控网络。我们进一步鉴定了这些细胞类型的特征过程、转录组谱和标记基因,揭示了双子叶和单子叶之间保守和分化的根发育途径。最后,我们利用时空模型从细胞特异性基因队列中鉴定了水稻根分生组织突变体,证明了该平台在功能遗传学研究方面的潜力。
二、研究思路
三、结果解读
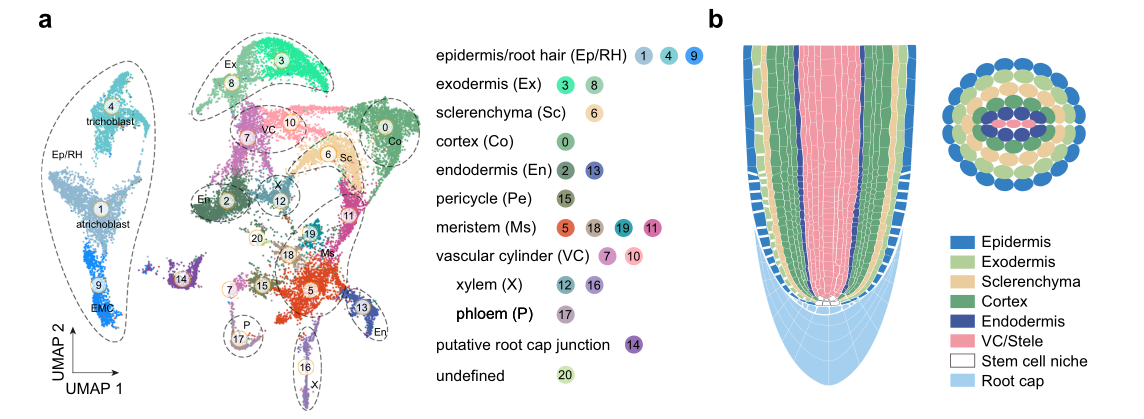
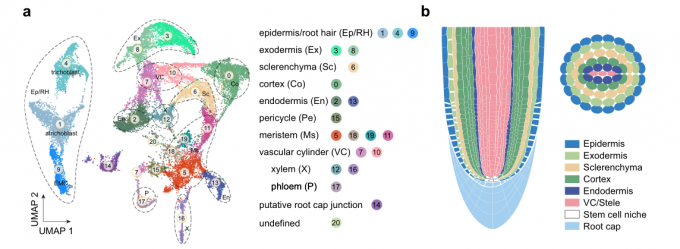
3.1 水稻根尖细胞图谱绘制
将野生型(ZH11)水稻幼苗的根(距根尖约1.0 cm, n= 90)采摘,消化成原生质体(无细胞壁的植物细胞),在商用10× Chromium平台上进行水稻根scRNA -seq测序,并且进行了两次生物学重复。最终,获得了一个包含27,469个过滤细胞与29,919个基因的基因表达矩阵。依据表达特征,所有细胞被划分为21个细胞类群。
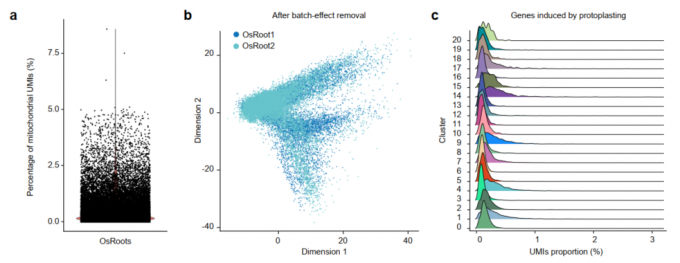
- 原生质体和线粒体基因对聚类的影响不大,以及去除批次效应后的结果
- 由于大多数细胞类型缺乏特定的标记,文中使用以下两种策略来注释水稻胚根细胞图谱中的细胞群体。首先,使用已经得到充分研究的水稻基因鉴定细胞群体类型。在UMAP上检测它们的表达模式有助于分配一些细胞簇。其次,作者对每个群体中特异性基因(超过30个基因)进行了RNA原位杂交分析。这些基因的表达模式不仅证实了上述注释,而且能够注释未知的细胞群体。
3.2 表皮细胞(Epidermal cells)的发育轨迹
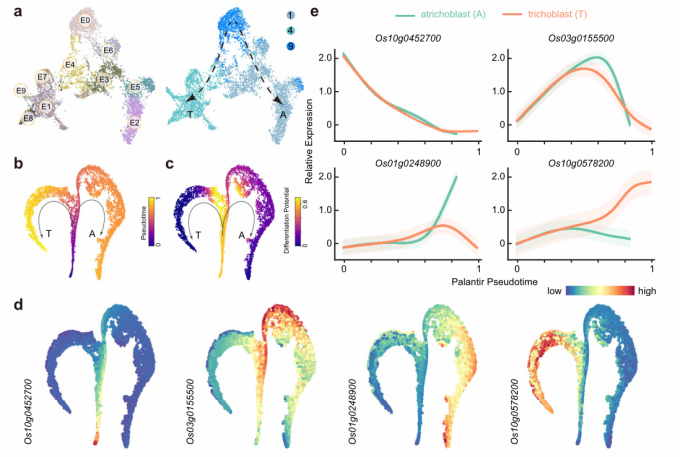
- 将epidermis/root hair population(clusters 1, 4, and 9)进行重新聚类,最终得到10个类群,从E0到E9。
- 从cluster 9开始,分叉形成两条轨迹,分别由cluster1和cluster4两个类群的细胞组成。作者使用Palantir进行拟时序分析,其结果与UMAP的聚类结果相似,推断的轨迹显示出从cluster 9逐渐过渡到早期、中期和晚期根毛细胞或到非根毛细胞。
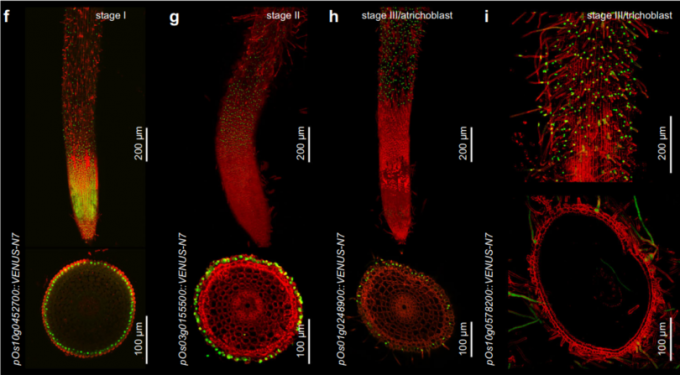
- 文中选择Os10g0452700、Os03g0155500、Os01g0248900和Os10g0578200作为cluster9、1和4的marker基因。其中,Os10g0452700的表达随着拟时间的延长而逐渐降低,接着是Os03g0155500的转录增加。Os01g0248900的表达在非根毛细胞发育后期升高,而Os10g0578200的表达仅在根毛细胞中逐渐升高。
- 荧光(Venus-N7)报告分析同样证实了由scRNA-seq数据集推断的表达模式。
总的来说,作者通过整合scRNA-seq数据、拟时序分析和报告基因分析等结果重建了水稻胚根根毛细胞和非根毛细胞发育过程中表皮细胞分化的过程。
3.3 基本组织细胞(Ground tissues)的发育轨迹
- 基本组织(fundamental tissue 或 ground tissue) 为1875年,J. Sachs 所提出植物体内的三种组织之一,另二者为表皮组织及维管束组织。
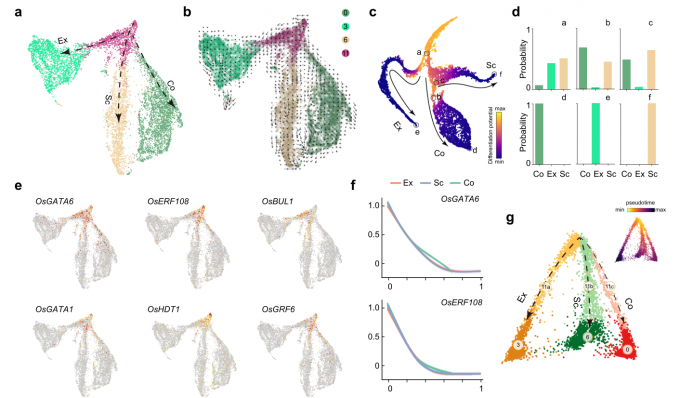
- 根据功能注释,聚类3、6和0分别被命名为外皮层、厚壁组织细胞层和皮层。作者通过仔细研究3D UMAP和t-SNE图上这三个类群的分布距离,发现这些细胞都与clsuter 11相连,因此,将cluster 11注释为分生组织细胞。因此,这一观察表明表明,cluster 11可能是基本组织共同的未分化祖细胞。
- 通过分析作者发现,在cluster 11中有多个转录因子高表达,例如OsGATA6和OsERF108的表达随着假时间逐渐减少。RNA原位杂交实验证实,OsGATA6和OsGRF6转录本主要聚集在RAM中心QC上方,并沿细胞分裂方向逐渐减少,直至达到分化区域。由此可见,cluster 11是由分生组织细胞组成,在水稻根尖形成了不同类型的基本组织和维管组织。
- 为了更好地了解基本组织分化背后的基因调控机制,作者将cluster 0,3,6,11重新聚类进行拟时序分析。结果显示出从分生组织细胞分化形成了外皮层、厚壁组织细胞层和皮层的过程。
- 以上分析表明OsGATA6在RAM中QC上方的中心位置表达,沿着细胞分裂方向其表达量逐渐降低,因此,OsGATA6可能在基本组织和维管组织的分化中起重要作用。为了验证这一假设,作者使用CRISPR-Cas9技术生成OsGATA6敲除突变体。纯合子Osgata6突变体植株具有矮化和短根的表型。而且,突变体的RAM较短,细胞分裂和分化出现缺陷。
3.4 通过ATAC-seq和scRNA-seq的整合分析推断细胞分化的转录调控机制
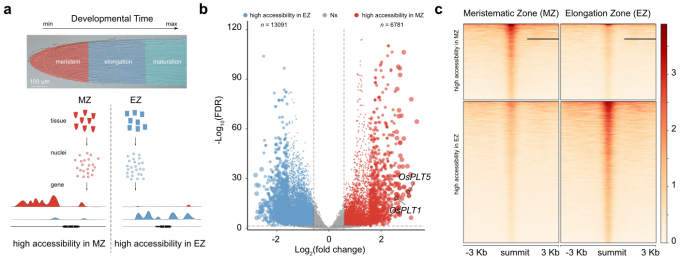
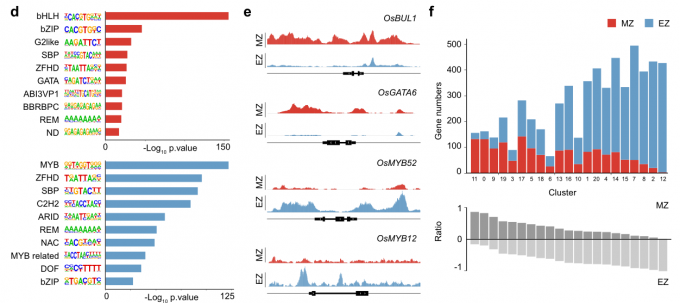
- 作者从根尖分生组织区(MZ)和伸长区(EZ)获取材料。对比MZ和EZ样品,发现染色质可及性发生了巨大变化。由此可知,在根尖分生组织细胞在分化过程中,染色质可及性放生了相应的变化。
- 作者使用HOMER软件鉴定了与染色质可及性变化相关的转录因子结合Motif。MZ和EZ样品富集了不同的转录因子结合Motif。转录因子家族如bHLH、bZIP和GATA的结合Motif在MZ中高度富集,而MYB转录因子结合Motif在EZ中富集。与这些结果一致的是,OsGATA6 (GATA转录因子)和OsBUL1(bHLH转录因子),这两个基因在cluster 11(对应于外皮层、厚壁组织细胞层和皮层细胞的分生组织细胞)中高表达,仅在MZ表现出开放的染色质状态。相比之下,MYB转录因子基因(cluster 12)MYB12和MYB52的调控区域在MZ区域是封闭的,而在EZ区域则是开放的。
- 为了检验在水稻根系发育过程中,给定基因的染色质可及性与其表达水平是否相关,作者对scRNA-seq和ATAC-seq数据集进行了综合分析。首先,将差异可及的基因分配到scRNA-seq实验揭示的细胞簇中。在MZ中,属于分生组织细胞的细胞群体(例如,cluster 9和cluster 11)显示了较高可及性的基因比例。相反,分配给分化组织的细胞群体(如cluster 2、7、8和12)显示出在EZ中拥有更高的染色质开放程度。
- cluster 9 (分生组织细胞)中的基因(Os10g0452700和OsEXPA11)在MZ中比在EZ中表现出更高的染色质开放程度。相比之下,在cluster 1和cluster4(分化的表皮细胞)中富集的基因(Os08g0112300, Os10g0490900, Os07g0176600和OsPKS16)在MZ中基本没有信号,但在EZ中却呈现出相反的信号趋势。
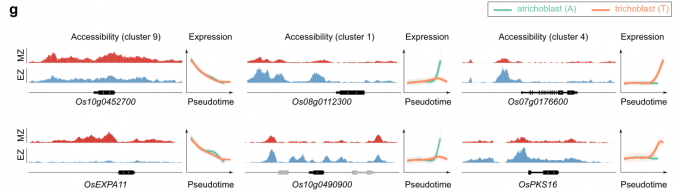
因此,上述结果表明,分析不同发育区域的染色质可及性可以揭示细胞分化的转录调控机制。
3.5 种间比较揭示了根系发育过程中的保守性和差异性
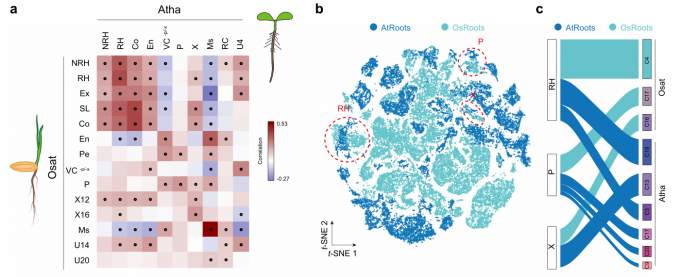
- 作者首先整合了3个已发表的拟南芥根scRNA-seq数据集,并重新进行了细胞聚类和分配,共鉴定出22个细胞簇。
- 通过对拟南芥和水稻根细胞簇的两两比较,发现分生组织、表皮细胞(无毛细胞和根毛细胞)、韧皮部和木质部细胞对应的细胞簇具有较高的相似性。有趣的是,水稻内胚层簇与拟南芥中相应簇的相关性较低,但与拟南芥中不含韧皮部和木质部细胞的维管组织(VC-P-X)呈正相关,表明这些细胞类型在功能上是一致的。水稻外皮层和拟南芥内皮层之间的相关性表明,它们是同源组织,与它们作为细胞运输屏障的功能一致。
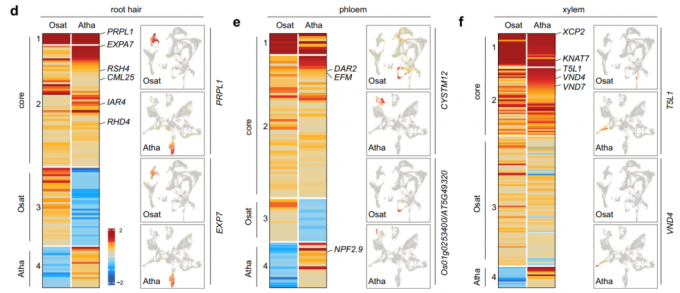
- 由于拟南芥和水稻在根毛、韧皮部和木质部上具有最高的一对一相似性,作者对它们进行了更详细的描述。对于根毛细胞类群,基因聚类分析显示了在两个物种的根毛中高表达的核心(保守)基因列表。值得注意的是,已知这些基因中的一些在根毛发育中发挥重要的调控作用。例如,PRPL1 (PROLINE-RICH PROTEIN-LIKE 1)和EXPA7 (EXPANSIN A7),它们都主要在根毛类群中表达。在拟南芥和水稻中有抑制根毛伸长的报道。同样,参与根毛发育的基因如RSH4、CML25 和RHD4也在两个物种共有的基因列表中。基因聚类分析同样观察到物种的特异性,表明每个物种的根毛具有独特的特性。与根毛类群的观察结果类似,作者还成功地鉴定出了与韧皮部和木质部细胞相对应的核心基因列表。
四、总结
- 作者通过聚类分群,在水稻胚根中公鉴定出21个细胞类群,分别由表皮、外皮层、厚壁细胞、皮层细胞、内皮层细胞、中柱鞘细胞、分生组织细胞、维管组织(韧皮部,木质部)、根冠细胞组成。
- 其次,通过对Cluster 1,4,9类群的进一步分析,探究了表皮细胞分化成根毛细胞与非根毛细胞的过程。
- 作者进一步通过对Cluster 0,3,6,11重新聚类分析。显示出从分生组织细胞分化形成了外皮层、厚壁组织细胞层和皮层的过程。
- 为了揭示细胞分化的转录调控机制,作者分析不同发育区域的染色质可及性
- 最后,作者探究了胚根细胞类型在单子叶-双子叶植物中的分裂与分化过程中的保守性。
单细胞文章解析(1):单细胞转录组图谱和染色质可及性揭示了水稻根的分化轨迹

